ņä£ļĪĀ
ļćīņĪĖņżæņØĆ ņŗĀĻ▓ĮņäĖĒżņØś ņåÉņāüņØä ņ£Āļ░£ĒĢśļ®░, ļåÆņØĆ ņ╣śņé¼ņ£©ņØä Ļ░Ćņ¦ĆļŖö ņ¦łĒÖśņØ┤ļŗż[1]. ļćīņĪĖņżæņØĆ Ēü¼Ļ▓ī ļćīĒśłĻ┤ĆņØ┤ Ēä░ņĀĖņä£ ļ░£ņāØĒĢśļŖö ņČ£Ēśłņä▒ ļćīņĪĖņżæĻ│╝ ļćīĒśłĻ┤ĆņØ┤ ļ¦ēĒśĆņä£ ļ░£ņāØĒĢśļŖö ĒŚłĒśłņä▒ ļćīņĪĖņżæņ£╝ļĪ£ ĻĄ¼ļČäļÉ£ļŗż[2]. ĒŚłĒśłņä▒ ļćīņĪĖņżæņØĆ ņé░ņåīĻ│ĄĻĖēņØś ļČĆņĪ▒ņ£╝ļĪ£ ņØĖĒĢ£ ĒÖ£ņä▒ņé░ņåīņóģ(reactive oxygen species)ņØś ņāØņä▒, adenosine triphosphate ņāØņä▒ Ļ░Éņåī, ņŚ╝ņ”Øļ░śņØæ ļ░Å ņäĖĒżņé¼ļ®ĖņØä ņ£ĀļÅäĒĢśņŚ¼ ņäĖĒż ņåÉņāüņØä ņØ╝ņ£╝Ēé©ļŗż[3,4]. ĒŚłĒśłņä▒ ļćīņĪĖņżæņŚÉ ņØśĒĢ£ ĻĖĆļŻ©ņĮöņŖż(glucose)ņØś Ļ░ÉņåīļŖö ņŗĀĻ▓ĮņäĖĒżņØś ņä▒ņןĻ│╝ ļČäĒÖöļź╝ ņ┤ēņ¦äņŗ£ĒéżļŖö ļŗ©ļ░▒ņ¦łņØś ĒÖ£ņä▒ņØä ņ¢ĄņĀ£ĒĢśņŚ¼ ņŗĀĻ▓ĮņäĖĒżņØś ņé¼ļ®ĖņØä ņ£ĀļÅäĒĢ£ļŗż[4]. ļśÉĒĢ£, ĒŚłĒśłņŚÉ ņØśĒĢ£ ĻĖĆļŻ©ĒāĆļ®öņØ┤ĒŖĖ(glutamate)ņØś Ļ│╝ļÅäĒĢ£ ņāØņä▒ņØĆ ņŗĀĻ▓ĮļÅģņä▒ņØä ņ£Āļ░£ĒĢśņŚ¼, ņé░ĒÖö ņŖżĒŖĖļĀłņŖżņŚÉ ņØśĒĢ£ ņŗĀĻ▓ĮņäĖĒżņØś ņåÉņāüņØä ņ£ĀļÅäĒĢ£ļŗż[5,6].
Chlorogenic acidļŖö Ļ│╝ņØ╝, ņ╗żĒö╝, ņ░©ņŚÉ ĒÆŹļČĆĒĢśĻ▓ī ņĪ┤ņ×¼ĒĢśļŖö ņ▓£ņŚ░ ĒÅ┤ļ”¼ĒÄśļåĆ ĒÖöĒĢ®ļ¼╝ ņżæ ĒĢśļéśņØ┤ļŗż[7]. ļŗżņ¢æĒĢ£ ņŗØļ¼╝ņŚÉņä£ ņČöņČ£ļÉ£ ĒÄśļåĆņé░ņØĆ ņ£ĀņÜ®ĒĢ£ ĻĖ░ļŖźņØä Ļ░Ćņ¦Ćļ®░, ĒŖ╣Ē׳ chlorogenic acidļŖö ĒĢŁņé░ĒÖö, ĒĢŁĻĘĀ, ĒĢŁļÅīņŚ░ļ│ĆņØ┤, ĒĢŁņŚ╝ņ”ØņØś ĒÜ©ļŖźņØä Ļ░Ćņ¦äļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[8]. Chlorogenic acidļŖö ņ¦Ćņ¦łļŗżļŗ╣ņ▓┤ņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ£ ņŚ╝ņ”Øļ░śņØæņØä ņÖäĒÖöņŗ£ĒéżĻ│Ā, ╬▓-amyloidņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ£ ĒĢ┤ļ¦ł(hippocampus)ņØś ņŗĀĻ▓ĮņäĖĒż ņé¼ļ®ĖņØä ņ¢ĄņĀ£ĒĢ£ļŗż[9,10]. ļśÉĒĢ£, chlorogenic acidļŖö ĒśłņĢĪ-ļćīņןļ▓Į(blood-brain barrier)ņØä ĒåĄĻ│╝ĒĢśļŖö ņןņĀÉņØä Ļ░ĆņĀĖ ņżæņČöņŗĀĻ▓ĮĻ│äņØś ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢśĻ│Ā, ņŗĀĻ▓ĮņäĖĒżņØś ņäĖĒżņé¼ļ®ĖņØä ņĀĆĒĢ┤ĒĢ£ļŗż[11]. ļśÉĒĢ£, ļćīņĪĖņżæ ļ¬©ļŹĖņŚÉņä£ chlorogenic acidļŖö ņé░ņåī-ĒżļÅäļŗ╣ Ļ▓░ĒĢŹ(oxygen-glucose deprivation)ņ£╝ļĪ£ļČĆĒä░ ņØ╝ņ░©ļē┤ļ¤░ņØä ļ│┤ĒśĖĒĢśĻ│Ā, ņŚ╝ņ”ØĻ│╝ ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżļź╝ Ļ░Éņåīņŗ£ņ╝£ ņŗĀĻ▓ĮņäĖĒżņØś ņé¼ļ®ĖņØä ļ¦ēļŖöļŗż[12]. ņĢ×ņäĀ ņŚ░ĻĄ¼ņŚÉņä£ ņÜ░ļ”¼ļŖö ņżæĻ░äļīĆļćīļÅÖļ¦źĒÅÉņćäņłĀ(middle cerebral artery occlusion, MCAO)ļĪ£ ņ£ĀļÅäļÉ£ ĒŚłĒśłņä▒ ļćīņĪĖņżæ ļ¬©ļŹĖņŚÉņä£ chlorogenic acidĻ░Ć ļīĆļćīņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ ņןņĢĀļź╝ Ļ░£ņäĀņŗ£ĒéżĻ│Ā, ļćīĻ▓ĮņāēĻ│╝ ņäĖĒżņé¼ļ®ĖņØś ņĀĢļÅäļź╝ ņÖäĒÖöņŗ£Ēé┤ņØä ļ│┤Ļ│ĀĒĢśņśĆļŗż[13].
EnolaseļŖö ņäĖĒżņØś ĒĢ┤ļŗ╣(glycolysis) Ļ│╝ņĀĢņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ĒĢ┤ļŗ╣ĒÜ©ņåī(glycolytic enzyme)ļĪ£ņä£, ņäĖĒżņØś ņä▒ņן ļ░Å ļČäĒÖöļź╝ ĒżĒĢ©ĒĢ£ ļŗżņ¢æĒĢ£ ņäĖĒż ĒÖ£ļÅÖņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗż[14]. EnolaseļŖö ╬▒, ╬▓, ╬│ņØś ņäĖĻ░Ćņ¦Ć ļÅÖņóģĒÜ©ņåīļź╝ Ļ░Ćņ¦Ćļ®░, ņØ┤ļōżņØĆ ņāØņ▓┤ ļé┤ ļČäĒż ņ£äņ╣śĻ░Ć ļŗżļź┤ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[15]. ╬▒-EnolaseļŖö ļīĆļČĆļČäņØś ņĪ░ņ¦üņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļ░śļ®┤, ╬▓-enolaseļŖö ĻĘ╝ņ£ĪņäĖĒżņŚÉņä£, ╬│-enolaseļŖö ņä▒ņłÖĒĢ£ ņŗĀĻ▓ĮņäĖĒżņÖĆ ņŗĀĻ▓Įļé┤ļČäļ╣äņäĖĒżņŚÉņä£ ļ░£ĒśäļÉ£ļŗż[16,17]. ĒŖ╣Ē׳, ╬│-enolaseļŖö ņŗĀĻ▓ĮĒŖ╣ņØ┤ņĀü(neurospecific) enolaseļĪ£ ļ¬ģļ¬ģļÉśļ®░, ņŗĀĻ▓ĮņäĖĒżņŚÉ ņśüņ¢æņØä Ļ│ĄĻĖēĒĢśĻ│Ā ņŗĀĻ▓ĮņĪ░ņ¦üņØä ļ│┤ĒśĖĒĢśļŖö ĒŖ╣ņä▒ņØä Ļ░Ćņ¦äļŗż[18,19]. ╬│-EnolaseļŖö phosphatidylinositol 3-kinase (PI3K)ņÖĆ mitogen-activated protein kinase (MAPK) ņŗĀĒśĖņĀäļŗ¼Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ņŗĀĻ▓ĮņäĖĒżņØś ņä▒ņן, ļČäĒÖö, ņ×¼ņāØņØä ņĪ░ņĀłĒĢśļ®░, ņŚ╝ņ”Øņä▒ ņé¼ņØ┤ĒåĀņ╣┤ņØĖņØä ņ┤ēņ¦äņŗ£ņ╝£ ņŗĀĻ▓Į ņŚ╝ņ”Øļ░śņØæņØä ņĪ░ņĀłĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[20,21]. ļśÉĒĢ£, ╬│-enolaseļŖö ņĀĢņāü ņĪ░ņ¦üņŚÉņä£ļŖö ņäĖĒż ļé┤ņŚÉ ņ׳ņ¦Ćļ¦ī ņäĖĒż ņåÉņāüņŚÉ ņØśĒĢ┤ ņäĖĒż ņÖĖ Ļ│ĄĻ░äņ£╝ļĪ£ ļ░®ņČ£ļÉśņ¢┤ ņåÉņāüņ£╝ļĪ£ļČĆĒä░ ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢ£ļŗż[22]. Chlorogenic acidļŖö ĒŚłĒśłņä▒ ļćīņåÉņāü ņŗ£, ņŗĀĻ▓ĮņäĖĒżņØś ņåÉņāüņØä ņÖäĒÖöņŗ£ņ╝£ ņŗĀĻ▓Įļ│┤ĒśĖ ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ņ¦Ćļ¦ī, chlorogenic acidņØś ╬│-enolase ļ░£Ēśä ņĪ░ņĀł ņŚ¼ļČĆņŚÉ Ļ┤ĆĒĢ£ ņŚ░ĻĄ¼ļŖö ņĢäņ¦üĻ╣īņ¦Ć ļ│┤Ļ│ĀļÉśņ¦Ć ņĢŖņĢśļŗż. ļö░ļØ╝ņä£, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö MCAOļĪ£ ņ£ĀļÅäļÉ£ ĒŚłĒśłņä▒ ļćīņåÉņāü ļ¬©ļŹĖņŚÉņä£ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ┤ ļ│ĆĒÖöĒĢśļŖö ļŗ©ļ░▒ņ¦ł ņżæ ╬│-enolase ļ░£ĒśäņØś ļ│ĆĒÖöļź╝ ņĪ░ņé¼ĒĢśĻ│Āņ×É ĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ņŗżĒŚśļÅÖļ¼╝ ļ░Å ņŗżĒŚśņäżĻ│ä
ņŗżĒŚśļÅÖļ¼╝ņØĆ (ņŻ╝)ņāśĒāĆņĮöļ░öņØ┤ņśżņĮöļ”¼ņĢä(Korea)ņŚÉņä£ ĻĄ¼ņ×ģĒĢ£ Spargue-Dawley ņłśņ╗Ę ĒØ░ņźÉ(7ņŻ╝ļĀ╣, 210-220 g, n = 64)ļĪ£, ĻĄ¼ņ×ģ Ēøä ņØ╝ņŻ╝ņØ╝ ļÅÖņĢł ņś©ļÅä(25Ōäā), ņŖĄļÅä(50% ┬▒ 5%), ļ╣ø(12 hours/12 hours ļ¬ģ/ņĢö)ņØ┤ ņĪ░ņĀłļÉśļŖö ļÅÖļ¼╝ņŗżņŚÉņä£ ņé¼ņ£ĪļÉśņŚłļŗż. ļ│Ė ņŗżĒŚśņØĆ phosphate buffered saline (PBS)ņØä Ēł¼ņŚ¼ĒĢ£ ņĀĢņāüĻĄ░(PBS + shamĻĄ░), chlorogenic acidļź╝ Ēł¼ņŚ¼ĒĢ£ ņĀĢņāüĻĄ░(chlorogenic acid + shamĻĄ░), PBSļź╝ Ēł¼ņŚ¼ĒĢ£ MCAOĻĄ░(PBS + MCAOĻĄ░), chlorogenic acidļź╝ Ēł¼ņŚ¼ĒĢ£ MCAOĻĄ░(chlorogenic acid + MCAOĻĄ░)ņ£╝ļĪ£ ļéśļłäņ¢┤ ņ¦äĒ¢ēļÉśņŚłļŗż. ļ¬©ļōĀ ļÅÖļ¼╝ņŗżĒŚś ņĀłņ░©ļŖö Ļ▓ĮņāüļīĆĒĢÖĻĄÉ ļÅÖļ¼╝ņŗżĒŚśņ£żļ”¼ņ£äņøÉĒÜī(Institutional Animal Care and Use Committee, IACUC)ņØś ņŖ╣ņØĖņØä ļ░øņĢä ņÜ┤ņśü ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ ņłśĒ¢ēļÉśņŚłļŗż(GNU-220222-R0021).
ņĢĮļ¼╝ Ēł¼ņŚ¼
ņÜ®ļ¦żņĀ£ļĪ£ņä£ PBSĻ░Ć ņé¼ņÜ®ļÉśņŚłĻ│Ā, chlorogenic acid (Sigma-Aldrich, USA)ļŖö PBSņŚÉ ļģ╣ņŚ¼ ņżĆļ╣äļÉśņŚłļŗż. ņŗżĒŚśļÅÖļ¼╝ļōżņØĆ MCAO ņłśņłĀ 2ņŗ£Ļ░ä ņĀäņŚÉ PBS ļśÉļŖö chlorogenic acidļź╝ ļ│ĄĻ░Ģ ņĢłņ£╝ļĪ£ ņŻ╝ņ×ģļÉśņŚłļŗż. Ēł¼ņŚ¼ļÉ£ chlorogenic acidņØś ņ¢æņØĆ 30 mg/kgņ£╝ļĪ£ ņĢ×ņä£ ļ│┤Ļ│ĀļÉ£ ļģ╝ļ¼ĖņØä ņ░ĖĻ│ĀĒĢśņŚ¼ Ļ▓░ņĀĢļÉśņŚłņ£╝ļ®░, chlorogenic acidļź╝ Ēł¼ņŚ¼ĒĢśņ¦Ć ņĢŖņØĆ ļÅÖļ¼╝ņØĆ PBSļ¦īņØä ļŗ©ļÅģ Ēł¼ņŚ¼ĒĢśņśĆļŗż[23]. ņŗżĒŚśĻĖ░Ļ░ä ļÅÖņĢł ļ¬©ļōĀ ļÅÖļ¼╝ļōżņØĆ ļ¼╝Ļ│╝ ņé¼ļŻīĻ░Ć ņ×Éņ£ĀļĪŁĻ▓ī Ļ│ĄĻĖēļÉśļŖö ĒÖśĻ▓ĮņŚÉņä£ ņé¼ņ£ĪļÉśņŚłļŗż.
ņżæĻ░äļīĆļćīļÅÖļ¦źĒÅÉņćäņłĀ
MCAOļŖö ĒŚłĒśłņä▒ ļīĆļćīņåÉņāüņØä ņ£ĀļÅäĒĢśĻĖ░ ņ£äĒĢ┤ ņŗżņŗ£ļÉśņŚłĻ│Ā ņĢ×ņä£ ļ│┤Ļ│ĀļÉ£ ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ņłśĒ¢ēļÉśņŚłļŗż[13]. ņŗżĒŚśļÅÖļ¼╝ņØĆ Zoletil (50 mg/kg; Virbac, France)ņØä ĻĘ╝ņ£Ī ļé┤ļĪ£ ņŻ╝ņ×ģĒĢśņŚ¼ ļ¦łņĘ©ļÉśņŚłĻ│Ā, ļ¬®ņØś ļ░░ņ¬Į ņĀĢņżæņäĀņØä ņĀłĻ░£ĒĢśņŚ¼ ņśżļźĖņ¬Į ņś©ļ¬®ļÅÖļ¦ź(common carotid artery)ņØä ļģĖņČ£ņŗ£ņ╝░ļŗż. ļģĖņČ£ņŗ£Ēé© ĒśłĻ┤ĆņØä ņŻ╝ļ│Ć ņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ĒĢ£ Ēøä, ņśżļźĖņ¬Į ļ░öĻ╣źļ¬®ļÅÖļ¦ź(exteral carotid artery), ņśżļźĖņ¬Į ņåŹļ¬®ļÅÖļ¦ź(internal carotid artery)ņØä ļģĖņČ£ņŗ£ņ╝░ļŗż. ņśżļźĖņ¬Į ļ░öĻ╣źļ¬®ļÅÖļ¦źņØś ļüØļČĆļČäņØä 6/0 ļ┤ēĒĢ®ņé¼ļĪ£ ļ¼ČĻ│Ā ņśżļźĖņ¬Į ņś©ļ¬®ļÅÖļ¦źņØĆ microvascular clipņØä ņØ┤ņÜ®ĒĢśņŚ¼ Ēśłļźśļź╝ ņ░©ļŗ©ĒĢ£ Ēøä, ņśżļźĖņ¬Į ļ░öĻ╣źļ¬®ļÅÖļ¦źņØä ņĀłņĀ£ĒĢśņśĆļŗż. ņśżļźĖņ¬Į ļ░öĻ╣źļ¬®ļÅÖļ¦źņØś ņĀłņĀ£ ļČĆņ£äļĪ£, Ļ░ĆņŚ┤ļĪ£ ļüØļČĆļČäņØä ļæźĻĖĆĻ▓ī ļ¦īļōĀ 4/0 nylon monofilamentļź╝ ņéĮņ×ģĒĢśņŚ¼ ņĢĮĻ░äņØś ņĀĆĒĢŁņØ┤ ļŖÉĻ╗┤ņ¦ł ļĢīĻ╣īņ¦Ć ņśżļźĖņ¬Į ņåŹļ¬®ļÅÖļ¦źņØä Ē¢źĒĢśņŚ¼ ļ░Ćņ¢┤ ļäŻņŚłļŗż. ņéĮņ×ģļÉ£ nylon monofilamentņØś ĻĖĖņØ┤ļŖö ņĢĮ 22-25 mmņśĆņ£╝ļ®░, filamentļŖö ņśżļźĖņ¬Į ņżæĻ░äļīĆļćīļÅÖļ¦źņØä ĒÅÉņćäĒĢśņŚ¼ ĒśłņĢĪņØś ĒØÉļ”äņØä ņ░©ļŗ©ņŗ£ņ╝░ļŗż. ņśżļźĖņ¬Į ņś©ļ¬®ļÅÖļ¦źņØä ņ░©ļŗ©Ē¢łļŹś microvascular clipņØä ņĪ░ņŗ¼ņŖżļ¤ĮĻ▓ī ņĀ£Ļ▒░ĒĢśņśĆļŗż. MCAOļź╝ ņŗżņŗ£ĒĢśņ¦Ć ņĢŖņØĆ shamĻĄ░ņØĆ nylon filament ņéĮņ×ģņØä ņĀ£ņÖĖĒĢśĻ│ĀļŖö MCAOĻĄ░Ļ│╝ ļÅÖņØ╝ĒĢ£ ļ░®ļ▓Ģņ£╝ļĪ£ ņ¦äĒ¢ēļÉśņŚłļŗż. ņŗżĒŚśļÅÖļ¼╝ņØĆ filament ņéĮņ×ģ 24ņŗ£Ļ░ä ĒøäņŚÉ ņĢłļØĮņé¼ļÉśņŚłĻ│Ā, ļīĆļćīĒö╝ņ¦ł ņĪ░ņ¦üņØä ņ▒äņĘ©ĒĢśņŚ¼ ņŗżĒŚśņĀäĻ╣īņ¦Ć -70ŌäāņŚÉņä£ ļ│┤Ļ┤ĆĒĢśņśĆļŗż.
ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ ņĀÉņłś Ļ▓Ćņé¼
ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ ņןņĢĀļŖö ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ ņĀÉņłś Ļ▓Ćņé¼(neurological deficit scoring test)ņŚÉ ņØśĒĢ┤ ĒÅēĻ░ĆļÉśņŚłļŗż[24]. MCAO ņłśņłĀ 24ņŗ£Ļ░ä ĒøäņŚÉ ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ ņĀÉņłś Ļ▓Ćņé¼ļź╝ ņŗżņŗ£Ļ│Ā, ņĀÉņłśļŖö 0-4ņĀÉņ£╝ļĪ£ ņĖĪņĀĢļÉśņŚłļŗż. ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉņØ┤ ņŚåļŖö ņĀĢņāüņĀüņØĖ ņ×ÉņäĖļŖö 0ņĀÉ(ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ ņŚåņØī), ļ░śļīĆņ¬Į ņĢ×ļŗżļ”¼ņØś ļČłņÖäņĀäĒĢ£ ņŚ░ņן ņ×ÉņäĖļŖö 1ņĀÉ(Ļ▓ĮļÅäņØś ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ), ļ░śļīĆņ¬Įņ£╝ļĪ£ ņ×Éļ░£ņĀüņ£╝ļĪ£ ņäĀĒÜīĒĢśļŖö Ē¢ēļÅÖņØĆ 2ņĀÉ(ņżæļÅäņØś ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ), ļ░śļīĆņ¬Įņ£╝ļĪ£ ļ¢©ņ¢┤ņ¦ĆĻ▒░ļéś ņ×ÉĻĘ╣ņŚÉ ļīĆĒĢ£ ļ»╝Ļ░ÉĒĢ£ ļ░śņØæ ļ░Å ļ░£ņ×æņØĆ 3ņĀÉ(ņŗ¼Ļ░üĒĢ£ ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ), ņ×Éļ░£ņĀü ņøĆņ¦üņ×äņØ┤ ņŚåĻ│Ā ņØśņŗØņØ┤ ņŚåļŖö ņāüĒā£ļŖö 4ņĀÉ(ļ¦żņÜ░ ņŗ¼Ļ░üĒĢ£ ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ)ņ£╝ļĪ£ ĻĄ¼ļČäļÉśņŚłļŗż.
ņĮöļäł ĒģīņŖżĒŖĖ
ņĮöļäł ĒģīņŖżĒŖĖ(corner test)ļŖö ņĢ×ņä£ ļ│┤Ļ│ĀļÉ£ ļ░®ļ▓Ģņ£╝ļĪ£ ņŗżņŗ£ļÉśņŚłļŗż[25]. ņŗżĒŚśļÅÖļ¼╝ņØĆ 30 ├Ś 20 ├Ś 1 cm3 Ēü¼ĻĖ░ņØś ļæÉ ĒīÉ ņé¼ņØ┤ņŚÉ ļåōņśĆļŗż. 30┬░ļĪ£ ļæÉ ĒīÉņØś Ļ░Ćņןņ×Éļ”¼ļź╝ ņŚ░Ļ▓░ĒĢśņśĆĻ│Ā, ņŗżĒŚśļÅÖļ¼╝ļōżņØä ļ¬©ņä£ļ”¼ļĪ£ ņØ┤ļÅÖņŗ£ņ╝░ļŗż. ņŗżĒŚśļÅÖļ¼╝ņØ┤ ĒīÉņØś ĻĄ¼ņäØņŚÉ ļÅäļŗ¼Ē¢łņØä ļĢī, ņ¢æņ¬Į ņĮ¦ņłśņŚ╝ņØĆ ļÅÖņŗ£ņŚÉ ĒīÉņŚÉ ļŗ┐ņĢśĻ│Ā, ļÅÖļ¼╝ļōżņØĆ ņŚ┤ļ”░ ļüØņØä Ē¢źĒĢśņŚ¼ ļÆżļĪ£ ļÅīņĢśļŗż. ļÅÖļ¼╝ļōżņØĆ MCAO ņłśņłĀ ņĀä 7ņØ╝ ļÅÖņĢł ĒøłļĀ©ņØä ļ░øņĢśĻ│Ā, ņśżļźĖņ¬ĮĻ│╝ ņÖ╝ņ¬Įņ£╝ļĪ£ ĒÜīņĀäĒĢśļŖö ļ╣äņ£©ņØ┤ ļ╣äņŖĘĒĢ£ ļÅÖļ¼╝ļ¦īņØä ļ│Ė ņŗżĒŚśņŚÉ ņé¼ņÜ®Ē¢łļŗż. ņĮöļäł ĒģīņŖżĒŖĖļŖö ņŗżĒŚśļÅÖļ¼╝ ļŗ╣ 10ĒÜīņö® ņĖĪņĀĢĒĢśņśĆņ£╝ļ®░ ņŗżĒŚśĻ▓░Ļ│╝ļŖö ņśżļźĖņ¬ĮĻ│╝ ņÖ╝ņ¬Į ĒÜīņĀä ĒܤņłśļĪ£ ļéśĒāĆļé┤ņŚłļŗż.
ņØ┤ņ░©ņøÉĻ▓ö ņĀäĻĖ░ņśüļÅÖ
ņśżļźĖņ¬Į ļīĆļćīĒö╝ņ¦ł ņĪ░ņ¦üņØä lysis buffer (8 M urea, 4% 3-[(3-Cholamidopropyl) dimethylammonium]-1-propanesulfonate [CHAPS], ampholytes and 40 mM Tris-HCl)ņŚÉ ļäŻņ¢┤ ĻĘĀņ¦łĒÖöĒĢ£ Ēøä, ņøÉņŗ¼ļČäļ”¼ĻĖ░(16,000g, 4Ōäā)ļĪ£ 20ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢśņŚ¼, ņāüņĖĄņĢĪņØä ĒÜīņłśĒĢśņśĆļŗż. ļŗ©ļ░▒ņ¦ł ņŗ£ļŻīļŖö Bradford ņŗ£ņĢĮ(Bio-Rad, USA)ņØä ņé¼ņÜ®ĒĢśņŚ¼ ņåīĒśłņ▓ŁņĢīļČĆļ»╝(bovine serum albumin)ņØä ĻĖ░ņżĆņ£╝ļĪ£ ņĀĢļ¤ēĒĢśņśĆņ£╝ļ®░, ņØ┤ņ░©ņøÉĻ▓ö ņĀäĻĖ░ņśüļÅÖ(2-dimensional gel electrophoresis)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļŗ©ļ░▒ņ¦ł ļČäņäØņØä ņ¦äĒ¢ēĒĢśņśĆļŗż. ļ©╝ņĀĆ, first-dimensional electrophoresisļź╝ ņłśĒ¢ēĒĢśĻĖ░ ņ£äĒĢ┤ immobilized pH gradient (IPG) gel strips (17 cm; Bio-Rad)ņŚÉ ļŗ©ļ░▒ņ¦ł ņŗ£ļŻī 50 ŃÄŹĻ│╝ sample buffer (8 M urea, 2% CHAPS, 20 mM dithiothreitol (DTT), 0.5% IPG buffer, bromophenol blue)ļź╝ ļäŻņ¢┤ 13ņŗ£Ļ░ä ļÅÖņĢł ņāüņś©ņŚÉņä£ ļ░śņØæņŗ£ņ╝░ļŗż. Ettan IPGphor 3 System (GE Healthcare, Sweden)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ stripsļź╝ 250 VļĪ£ 15ļČä, 10,000 VļĪ£ 3ņŗ£Ļ░ä, 10,000 VņŚÉņä£ 50,000 VļĪ£ ņāüņŖ╣ĒĢĀ ļĢīĻ╣īņ¦Ć focusingņØä ņ¦äĒ¢ēĒĢ£ Ēøä, 1% DTTĻ░Ć ĒżĒĢ©ļÉ£ equilibration buffer (6 M urea, 30% glycerol, 2% sodium dodecyl sulfate (SDS), 50 mM Tris-HCl [pH 8.8])ņŚÉņä£ 15ļČä, 2.5% iodoacetamideĻ░Ć ĒżĒĢ©ļÉ£ equilibration bufferņŚÉņä£ 15ļČä ļ░śņØæņŗ£ņ╝░ļŗż. StipņØĆ SDS-polyacrylamide gradient gels (7.5%-17.5%)ņŚÉ ņś¼ļĀż ņØ┤ņ░©ņøÉĻ▓ö ņĀäĻĖ░ņśüļÅÖņØ┤ ņ¦äĒ¢ēļÉśņŚłĻ│Ā, bromophenol blue dyeĻ░Ć gelņØś ļ░öļŗźņŚÉ ļÅäļŗ¼ĒĢĀ ļĢīĻ╣īņ¦Ć Protein-ŌģĪ Ōģ¬ electrophoresis equipment (Bio-Rad)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 10ŌäāņŚÉņä£ 5 mAļĪ£ 2ņŗ£Ļ░ä, 10 mAļĪ£ 10ņŗ£Ļ░ä ņĀäĻĖ░ņśüļÅÖļÉśņŚłļŗż. GelņØĆ Ļ│ĀņĀĢņĢĪ(12% acetic acid and 50% methanol)ņŚÉņä£ 2ņŗ£Ļ░ä Ļ│ĀņĀĢļÉśņŚłĻ│Ā, 50% ethanolņŚÉ 20ļČäĻ░ä ļ░śņØæņŗ£ņ╝░ļŗż. GelņØĆ silver solution (0.2% silver nitrate)ņŚÉņä£ 20ļČä ņŚ╝ņāēļÉśņŚłĻ│Ā, ĒśäņāüņĢĪ(0.2% sodium carbonate)ņ£╝ļĪ£ Ēśäņāü ļÉ£ Ēøä Agfar ARCUS 1200 ņŖżņ║Éļäł(Agfa-Gevaert, Belgium)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņØ┤ļ»Ėņ¦Ć ĒīīņØ╝ļĪ£ ņĀĆņןļÉśņŚłļŗż. ņØ┤ļ»Ėņ¦ĆļŖö PDQuest 2-D analysis software (Bio-Rad)ļĪ£ ļČäņäØļÉśņŚłĻ│Ā, Ēæ£ņĀü ļŗ©ļ░▒ņ¦ł spotņØä ļ¢╝ņ¢┤ļé┤ņ¢┤, trypsinņØ┤ ĒĢ©ņ£ĀĒĢ£ ņÖäņČ®ņĢĪņŚÉ ļ░śņØæņŗ£Ēé© Ēøä matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF)ļź╝ ņ¦äĒ¢ēĒĢśņśĆļŗż. ņČöņČ£ĒĢ£ ļŗ©ļ░▒ņ¦łņŚÉ ļīĆĒĢśņŚ¼ Voyager System DE-STR MALDI-TOF mass spectrometer (Life Technologies, Carlsbad, CA, USA)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ mass spectrometryļź╝ ņŗżņŗ£ĒĢśņśĆĻ│Ā, MS-FITĻ│╝ ProFound programņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļŗ©ļ░▒ņ¦łņØś ņĀĢļ│┤ļź╝ ņČöņĀüĒĢśĻ│Ā, NCBI databaseļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļŗ©ļ░▒ņ¦ł ņä£ņŚ┤ņØä ĒÖĢņØĖĒĢśņśĆļŗż.
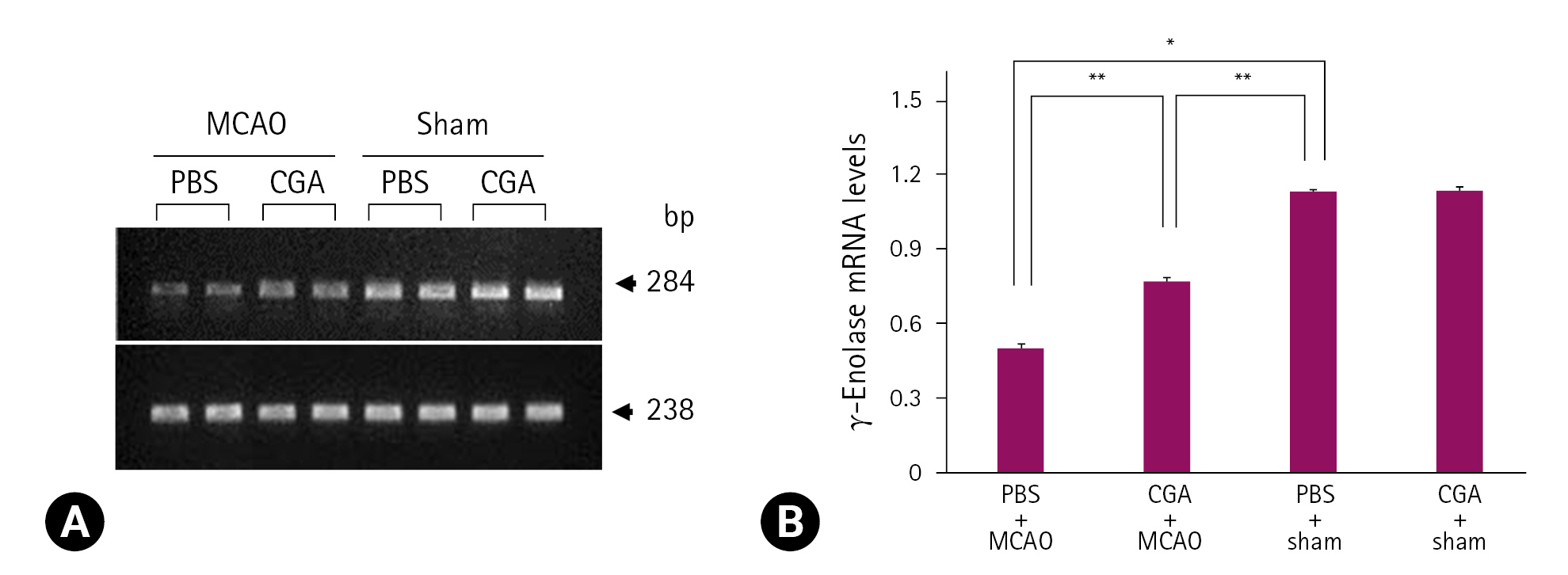
Reverse transcription- polymerase chain reaction
ņśżļźĖņ¬Į ļīĆļćīĒö╝ņ¦ł ņĪ░ņ¦üņØĆ Trizol Reagent (Life Technologies, USA)ņŚÉ ļäŻņ¢┤ ĻĘĀņ¦łĒÖöļÉśņŚłĻ│Ā, ņøÉņŗ¼ļČäļ”¼ĻĖ░(13,000g, 4Ōäā)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 20ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢ£ Ēøä ņāüņĖĪņĢĪņŚÉņä£ total RNAļź╝ ņČöņČ£ĒĢśņśĆļŗż. Reverse transcription-polymerase chain reaction (PCR)ņØĆ superscript III firststrand system (Invitrogen, USA)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņłśĒ¢ēļÉśņŚłĻ│Ā, ļ░®ļ▓ĢņØĆ ņĀ£ņĪ░ņé¼ņØś ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ total DNAļź╝ ņŚŁņĀäņé¼ņŗ£ņ╝£ ļŗ©ņØ╝ Ļ░Ćļŗź ņāüļ│┤ņĀü DNAļź╝ ņ¢╗ņŚłĻ│Ā, PCRņØä ĒåĄĒĢ┤ cDNAņØś ╬│-enolaseņØś ņ£ĀņĀäņ×É ņä£ņŚ┤ņØä ņ”ØĒÅŁņŗ£ņ╝░ļŗż. ņé¼ņÜ®ļÉ£ ╬│-enolaseņØś primer ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ 5'-TGGATCTCCATACTG CCAAAG-3' (forward)Ļ│╝ 5'-CCAACTCCTCTTCAA TCCTCAT-3' (reverse)ņśĆļŗż. ╬▓-actinņØś primer ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ 5ŌĆÖ-GGGTCAGAAGGACTCCTACG-3ŌĆÖ(forward)Ļ│╝ 5ŌĆÖ-GGTCTCAAACATGATCTGGG-3ŌĆÖ(reverse)ņśĆļŗż. PCR ļ░śņØæņØĆ 94ŌäāņŚÉņä£ 5ļČä denature Ļ│╝ņĀĢ, 94ŌäāņŚÉņä£ 30ņ┤ł, 54ŌäāņŚÉņä£ 30ņ┤ł, 72ŌäāņŚÉņä£ 1ļČäņØś 30 cycles Ļ│╝ņĀĢņØä Ļ▒░ņ│żņ£╝ļ®░, ņØ┤Ēøä 72ŌäāņŚÉņä£ 10ļČä enlongation Ļ│╝ņĀĢņ£╝ļĪ£ ņŗ£Ē¢ēļÉśņŚłļŗż. PCR productsļŖö Loading STAR (Dyne Bio, Korea)ņÖĆ Ēś╝ĒĢ®ĒĢśņŚ¼ 1% agarose gelņŚÉņä£ ņĀäĻĖ░ņśüļÅÖļÉśņŚłĻ│Ā, ļČäļ”¼ļÉ£ PCR product bandsļź╝ ultraviolet-lightņŚÉņä£ Ļ┤Ćņ░░ĒĢ£ Ēøä, SigmaGel 1.0 (Jandel Scientific, USA)ņÖĆ SigmaPlot 4.0 (SPSS Inc., USA) ĒöäļĪ£ĻĘĖļשņ£╝ļĪ£ ļČäņäØļÉśņŚłļŗż. ╬│-Enolase ļ░£Ēśä ņłśņżĆņØĆ ╬▓-actinņØś ļ░£Ēśä Ļ░ĢļÅäņŚÉ ļīĆĒĢ£ ╬│-enolaseņØś Ļ░ĢļÅäļĪ£ ĒÅēĻ░ĆļÉśņŚłļŗż.
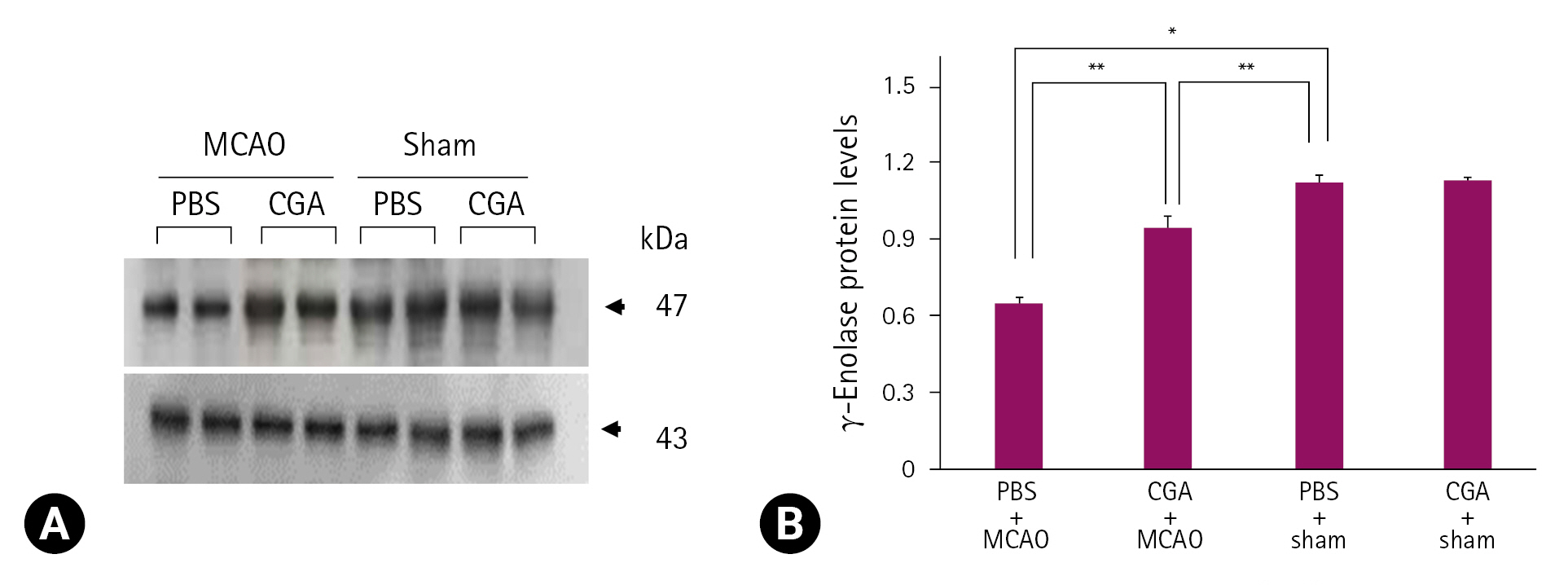
Western blot ļČäņäØ
ņśżļźĖņ¬Į ļīĆļćīĒö╝ņ¦ł ņĪ░ņ¦üņØĆ lysis buffer (1% Triton X-100 and 1 mM EDTA in phosphate buffer saline)ņŚÉ ļäŻņ¢┤ ļČäņćäĻĖ░(homogenizer)ļĪ£ ļČäņćäļÉśņŚłĻ│Ā, ļČäņćäļÉ£ Ēś╝ĒĢ®ļ¼╝ņØĆ ņøÉņŗ¼ļČäļ”¼ĻĖ░(15,000g, 4Ōäā)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 20ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢ£ Ēøä ņāüņĖĄņĢĪņØĆ ĒÜīņłśļÉśņŚłļŗż. ļŗ©ļ░▒ņ¦ł ņĖĪņĀĢņØĆ bicinchoninic acid protein assay kit (Pierce, USA)ļź╝ ņØ┤ņÜ®ĒĢśņśĆĻ│Ā, ņåīĒśłņ▓ŁņĢīļČĆļ»╝ņØä ĻĖ░ņżĆņ£╝ļĪ£ ļŗ©ļ░▒ņ¦ł ņŗ£ļŻīņØś ņĀĢļ¤ēņØ┤ ņØ┤ļŻ©ņ¢┤ņĪīļŗż. SDS-polyacrylamide gelņŚÉ ļŗ©ļ░▒ņ¦ł ņŗ£ļŻī(30 ŃÄŹ)ļź╝ ļäŻņ¢┤ ņĀäĻĖ░ņśüļÅÖĒĢ£ Ēøä, ļŗ©ļ░▒ņ¦łļōżņØĆ gelļĪ£ļČĆĒä░ poly-vinylidene fluoride membrane (Millipore, USA)ņ£╝ļĪ£ ņś«Ļ▓©ņĪīļŗż. ļ╣äĒŖ╣ņØ┤ņĀüņØĖ ĒĢŁņ▓┤ļ░śņØæņØä ņ¢ĄņĀ£ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ 5% skim milk ņÜ®ņĢĪņŚÉ membraneņØĆ 1ņŗ£Ļ░ä ļÅÖņĢł ļ░śņØæļÉśņŚłĻ│Ā, tris-buffered saline with 0.1% Tween 20 (TBST)ļĪ£ 5ļČäĻ░ä 3ĒÜī ņäĖņ▓ÖļÉśņŚłļŗż. 1ņ░© ĒĢŁņ▓┤ļĪ£ņä£ ╬│-enolase (1:1,000; Santa Cruz Biotechnology, USA) ļśÉļŖö ╬▓-actin (1:1,000; Santa Cruz Biotechnology)ņØä 1ņŗ£Ļ░ä ļÅÖņĢł ļ░śņØæņŗ£ņ╝░Ļ│Ā, TBSTļĪ£ 3ĒÜī ņäĖņ▓ÖĒĢ£ Ēøä, horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5,000, Pierce)ļź╝ 2ņ░© ĒĢŁņ▓┤ļĪ£ ņ▓śļ”¼ĒĢśņśĆļŗż. TBSTļĪ£ ņäĖņ▓ÖĒĢ£ Ēøä membraneņØä enhanced chemiluminescence Western blot analysis system (Amersham Pharmacia Biotech, USA)ņŚÉ 1ļČäĻ░ä ļ░śņØæņŗ£Ēé© Ēøä, X-ray filmņŚÉ 1ļČäĻ░ä ļģĖņČ£ņŗ£ņ╝░Ļ│Ā, ĒśäņāüņĢĪņŚÉņä£ ĒśäņāüĒĢ£ Ēøä Ļ│ĀņĀĢņĢĪņ£╝ļĪ£ Ļ│ĀņĀĢĒĢśņśĆļŗż. SigmaGel 1.0Ļ│╝ SigmaPlot 4.0ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļ░£ņāēņØä ĒÖĢņØĖĒĢśĻ│Ā ļŗ©ļ░▒ņ¦ł ļ░£Ēśäņ¢æņØä ņĖĪņĀĢĒĢśņśĆļŗż. ╬│-Enolase ļ░£Ēśä ņłśņżĆņØĆ ╬▓-actinņØś ļ░£Ēśä Ļ░ĢļÅäņŚÉ ļīĆĒĢ£ ╬│-enolaseņØś Ļ░ĢļÅäļĪ£ ĒÅēĻ░ĆļÉśņŚłļŗż.
ļ®┤ņŚŁĒśĢĻ┤æņŚ╝ņāē
ņĀüņČ£ļÉ£ ļćīņĪ░ņ¦üņØä 4% ņÖäņČ® Ēżļź┤ļ¦Éļ”░ Ļ│ĀņĀĢņĢĪņŚÉņä£ 24ņŗ£Ļ░ä Ļ│ĀņĀĢņŗ£ņ╝░ļŗż. Ļ│ĀņĀĢļÉ£ ņĪ░ņ¦üņØĆ ņłśņäĖ Ēøä ethanol (70%-100%)Ļ│╝ xyleneņ£╝ļĪ£ ņłśņäĖ ļ░Å Ēł¼ļ¬ģĒÖö Ļ│╝ņĀĢņØä Ļ▒░ņ│żļŗż. ĒīīļØ╝ĒĢĆ Ēżļ¦żĻĖ░(paraffin embedding center; Leica, Germany)ņŚÉņä£ Ēżļ¦żĒĢ£ Ēøä, ĒÜīņĀäņŗØ ļ░ĢņĀłĻĖ░(Leica)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 4 ╬╝mņØś ļæÉĻ╗śļĪ£ ļ░ĢņĀłĒĢśņśĆļŗż. ļ░ĢņĀłļÉ£ ņĪ░ņ¦üņØĆ ņŖ¼ļØ╝ņØ┤ļō£ĻĖĆļØ╝ņŖż ņ£äņŚÉ ņś¼ļĀżņĪīĻ│Ā, ņŗĀņĀäĻĖ░(slide warmer; Thermo Fisher Scientific, USA)ņŚÉņä£ Ļ▒┤ņĪ░ļÉśņŚłļŗż. ļ░ĢņĀłļÉ£ ņĪ░ņ¦üņØĆ xyleneņ£╝ļĪ£ ĒāłĒīīļØ╝ĒĢĆļÉśņŚłĻ│Ā, ethanol (100%-70%)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ĒĢ©ņłśĒÖö Ļ│╝ņĀĢņØä Ļ▒░ņ│żļŗż. ņłśņäĖļÉ£ ņĪ░ņ¦üņØĆ ĒĢŁņøÉņØä ļģĖņČ£ņŗ£ĒéżĻĖ░ ņ£äĒĢ┤ ņĀäņ×ÉļĀłņØĖņ¦Ćļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 0.1 M sodium citrate buffer (pH 6.0)ņŚÉ ļ░śņØæņŗ£ņ╝░Ļ│Ā, ļ╣äĒŖ╣ņØ┤ņĀüņØĖ ĒĢŁņ▓┤ņØś Ļ▓░ĒĢ®ņØä ņ¢ĄņĀ£ĒĢśĻĖ░ ņ£äĒĢ┤ 0.5% fetal bovine serumņŚÉ 1ņŗ£Ļ░ä ņ▓śļ”¼ļÉśņŚłļŗż. 1ņ░© ĒĢŁņ▓┤ļĪ£ anti-╬│-enolase (1:100; Santa Cruz Biotechnology)ņØä 4ŌäāņŚÉņä£ 12ņŗ£Ļ░ä ļ░śņØæņŗ£ņ╝░Ļ│Ā, PBSļĪ£ 3ĒÜī ņäĖņ▓ÖĒĢ£ Ēøä, 2ņ░© ĒĢŁņ▓┤ļĪ£ņä£ fluorescein isothiocyanate-conjugated secondary antibody (1:100; Santa Cruz Biotechnology)ļź╝ ņŗżņś©ņŚÉņä£ 1ņŗ£Ļ░ä ļ░śņØæņŗ£ņ╝░ļŗż. PBSļĪ£ ņäĖņ▓Ö Ēøä, ĒĢĄņØĆ 4',6-diamidino-2-phenylindole (DAPI; Santa Cruz Biotechnology)ļĪ£ ņŚ╝ņāēļÉśņŚłĻ│Ā, ņłśņäĖ Ēøä Ultra Cruz mounting mediumņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņĪ░ņ¦üņØä ļ┤ēņ×ģĒĢśņśĆļŗż. Ļ│Ąņ┤łņĀÉ Ēśäļ»ĖĻ▓Į(confocal microscope, FV- 1000; Olympus, Japan)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņŚ╝ņāēļÉ£ ļČĆņ£äļź╝ ĒÖĢņØĖĒĢśņśĆĻ│Ā, Ļ▓░Ļ│╝ ļČäņäØņØä ņ£äĒĢśņŚ¼ ņśżļźĖņ¬Į ļīĆļćīĒö╝ņ¦łņØś 5Ļ░£ ļČĆņ£äļź╝ ļ¼┤ņ×æņ£äļĪ£ ņäĀĒāØĒĢśņśĆļŗż. ļīĆļćīĒö╝ņ¦łņØś ņØ┤ļ»Ėņ¦ĆĻ░Ć ņ║Īņ▓śļÉśņŚłĻ│Ā, Image-Pro Plus image analysis softwareļĪ£ ņāüļīĆņĀüņØĖ ĒåĄĒĢ® ļ░ĆļÅä(relative integrated density)ļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ╬│-Enolase ļ░£Ēśä ņłśņżĆņØĆ PBS + shamĻĄ░ņØś ĒåĄĒĢ® Ļ░ĢļÅäņŚÉ ļīĆĒĢ£ Ļ░ü ņŗżĒŚśĻĄ░ņØś ĒåĄĒĢ® Ļ░ĢļÅäņØś ļ╣äņ£©ļĪ£ ļéśĒāĆļé┤ņŚłļŗż.
Ļ▓░Ļ│╝
MCAO ņåÉņāü ņŗ£ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ņŗĀĻ▓ĮĒ¢ēļÅÖĒĢÖņĀü ņןņĢĀ Ļ░£ņäĀ ĒÜ©Ļ│╝
ļ©╝ņĀĆ ╬│-enolase ļ░£ĒśäņØś ļ│ĆĒÖöļź╝ ņĪ░ņé¼ĒĢśĻĖ░ņĀäņŚÉ MCAO ņåÉņāü ņŗ£ chlorogenic acidņØś ņŗĀĻ▓ĮĒ¢ēļÅÖĒĢÖņĀü ņןņĢĀņØś Ļ░£ņäĀņØä ĒÖĢņØĖĒĢśņśĆļŗż. MCAO ņåÉņāüņØĆ ņŗ¼Ļ░üĒĢ£ ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ ņןņĢĀļź╝ ļéśĒāĆļé┤ņŚłļŗż. PBS Ēł¼ņŚ¼ĻĄ░ņŚÉņä£ļŖö ĒĢ£ ļ░®Ē¢źņ£╝ļĪ£ ĒÜīņĀä, ņøĆņ¦üņ×ä ļČĆņĪ▒, ļ░£ņ×æ ļō▒ ņŗ¼Ļ░üĒĢ£ Ē¢ēļÅÖ ņןņĢĀļź╝ ļ│┤ņśĆņ£╝ļéś, chlorogenic acidņØś Ēł¼ņŚ¼ļŖö ņØ┤ļ¤¼ĒĢ£ ņןņĢĀļź╝ Ļ░£ņäĀņŗ£ņ╝░ļŗż. ShamĻĄ░ņŚÉņä£ļŖö PBSņÖĆ chlorogenic acidņØś Ēł¼ņŚ¼ņÖĆ ņāüĻ┤ĆņŚåņØ┤ Ē¢ēļÅÖ ņןņĢĀ ņ”ØņāüņØ┤ ļéśĒāĆļéśņ¦Ć ņĢŖņĢśļŗż(Table 1). Ļ░ÉĻ░üņÜ┤ļÅÖņØś ļ░®Ē¢ź Ēī©Ēä┤ņØä ĒÅēĻ░ĆĒĢśĻĖ░ ņ£äĒĢ┤ ņĮöļäł ĒģīņŖżĒŖĖļź╝ ņłśĒ¢ēĒĢśņśĆĻ│Ā, MCAO ņłśņłĀĻĄ░ņØĆ ņśżļźĖņ¬Į ļćīņåÉņāüņ£╝ļĪ£ ņØĖĒĢśņŚ¼ ņśżļźĖņ¬Įņ£╝ļĪ£ļ¦ī ļ░śņØæņØä ļ│┤ņØ┤ļŖö Ē¢ēļÅÖ ņןņĢĀļź╝ ļéśĒāĆļāłņŚłĻ│Ā, chlorogenic acidņØś Ēł¼ņŚ¼ļŖö ņØ┤ļ¤¼ĒĢ£ ļ░śņØæņØä ņÖäĒÖöņŗ£ņ╝░ļŗż(Table 1). ļ│Ė ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ ĒģīņŖżĒŖĖļŖö MCAOļź╝ ĒåĄĒĢ£ ĒŚłĒśłņä▒ ļćīņĪĖņżæ ļÅÖļ¼╝ļ¬©ļŹĖņØ┤ ņל ĒÖĢļ”ĮņØ┤ ļÉśņŚłļŖöņ¦Ćļź╝ ĒÖĢņØĖĒĢśĻ│Āņ×É ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ Ļ▓Ćņé¼ļź╝ ņŗżņŗ£ĒĢśņśĆĻ│Ā, chlorogenic acidņŚÉ ņØśĒĢ£ ņŗĀĻ▓Įļ│┤ĒśĖ ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśļŖö ĻĖ░ļ│Ė ņ×ÉļŻīļź╝ ņĀ£ņŗ£ĒĢśĻ│Āņ×É, Ļ▓░Ļ│╝ ļČĆļČäņŚÉ ņéĮņ×ģĒĢśņśĆļŗż.
MCAO ņåÉņāü ņŗ£ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolase ļ░£Ēśä Ļ░ÉņåīņØś ņÖäĒÖö
ĒöäļĪ£Ēģīņśżļ»╣ņŖż ļČäņäØļ▓ĢņØä ĻĖ░ļ░śņ£╝ļĪ£ MCAOņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉ£ ļīĆļćīĒŚłĒśł ņŗ£ņŚÉ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolase ļŗ©ļ░▒ņ¦łņØś ļ░£Ēśä ļ│ĆĒÖöļź╝ Ļ┤Ćņ░░ĒĢśņśĆļŗż. ╬│-EnolaseņØś peptide massļŖö 14/70ņØ┤ņŚłĻ│Ā sequence coverageļŖö 34%ņśĆļŗż. MCAO ņłśņłĀņØĆ ļīĆļćīĒö╝ņ¦łņØś ╬│-enolase ļŗ©ļ░▒ņ¦ł ļ░£Ēśä Ļ░Éņåīļź╝ ņ£ĀļÅäĒĢśņśĆĻ│Ā, MCAOņŚÉ ņØśĒĢ£ ╬│-enolase ļ░£ĒśäņØś Ļ░ÉņåīļŖö chlorogenic acidņØś Ēł¼ņŚ¼ļĪ£ ņÖäĒÖöļÉśņŚłļŗż(Fig. 1A). ĻĘĖļ¤¼ļéś ņłśņłĀĻ│╝ņĀĢņŚÉņä£ nylonņØä ņéĮņ×ģĒĢśņ¦Ć ņĢŖņØĆ shamĻĄ░ņŚÉņä£ļŖö PBSņÖĆ chlorogenic acid Ēł¼ņŚ¼ņÖĆ ņāüĻ┤ĆņŚåņØ┤ ╬│-enolaseņØś ļ░£ĒśäņØĆ ņ£Āņé¼ĒĢśĻ▓ī ļéśĒāĆļé¼ļŗż. ╬│-Enolase ļ░£Ēśä ņĀĢļÅäļŖö PBS + shamĻĄ░ņØś Ļ▓░Ļ│╝ņŚÉ ļīĆĒĢ£ ļ╣äņ£©ļĪ£ ĒÅēĻ░ĆļÉśņŚłĻ│Ā, PBS + MCAOĻĄ░ņŚÉņä£ļŖö 0.48 ┬▒ 0.03ņØ┤ņŚłĻ│Ā, chlorogenic acid + MCAOĻĄ░ņŚÉņä£ļŖö 0.93 ┬▒ 0.04ņśĆļŗż(Fig. 1B). Reverse transcription-PCRĻ│╝ Western blot ļČäņäØņØĆ chlorogenic acid Ēł¼ņŚ¼Ļ░Ć MCAO ņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ ╬│-enolase ļ░£ĒśäņØś Ļ░Éņåīļź╝ ņÖäĒÖöņŗ£Ēé┤ņØä ļ│┤ņŚ¼ņŻ╝ņŚłļŗż(Figs. 2, 3). ╬│-Enolase mRNA ļ░£ĒśäņØĆ MCAO ņåÉņāüņ£╝ļĪ£ Ļ░ÉņåīļÉśņŚłĻ│Ā, chlorogenic acid Ēł¼ņŚ¼ļŖö ņØ┤ļōż Ļ░Éņåīļź╝ ņÖäĒÖöņŗ£ņ╝░ļŗż(Fig. 2A). ╬│-Enolase mRNA ļ░£ĒśäņØĆ PBS ļśÉļŖö chlorogenic acidņØä ņ▓śļ”¼ĒĢ£ shamĻĄ░ņŚÉņä£ļŖö ņ£Āņé¼ĒĢśĻ▓ī ļéśĒāĆļé¼ņ£╝ļ®░, PBS + MCAOĻĄ░ņŚÉņä£ļŖö 0.50 ┬▒ 0.02ņśĆĻ│Ā, chlorogenic acid + MCAOĻĄ░ņŚÉņä£ļŖö 0.77 ┬▒ 0.02ņśĆļŗż(Fig. 2B). ╬│-Enolase ļŗ©ļ░▒ņ¦łņØś ļ░£ĒśäņØĆ PBS + MCAOĻĄ░Ļ│╝ chlorogenic acid + MCAOĻĄ░ņŚÉņä£ Ļ░üĻ░ü 0.66 ┬▒ 0.03Ļ│╝ 0.95 ┬▒ 0.04ļĪ£ ļéśĒāĆļé¼ļŗż(Fig. 3A and B). ļ®┤ņŚŁĒśĢĻ┤æņŚ╝ņāē(immunofluorescence stain) Ļ▓░Ļ│╝ļŖö MCAO ņåÉņāü ņŗ£ņŚÉ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolase ļ░£ĒśäņØś ļ│ĆĒÖöļź╝ ļ│┤ņŚ¼ņŻ╝ņŚłļŗż(Fig. 4A). ļīĆļćīĒö╝ņ¦ł ļČĆņ£äņØś ╬│-enolaseņØś Ļ░ĢļÅäļŖö shamĻĄ░ņŚÉ ļ╣äĒĢ┤ PBS + MCAOĻĄ░ņŚÉņä£ļŖö ņ£ĀņØśĒĢśĻ▓ī Ļ░ÉņåīĒĢśņśĆĻ│Ā, chlorogenic acid + MCAOĻĄ░ņŚÉņä£ļŖö MCAO ņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ Ļ░Éņåīļź╝ ņÖäĒÖöņŗ£ņ╝░ļŗż(Fig. 4A). ļ®┤ņŚŁĒśĢĻ┤æņŚ╝ņāēņØä ĒåĄĒĢ£ ╬│-enolase ļ░£ĒśäņØĆ PBS + MCAOĻĄ░Ļ│╝ chlorogenic acid + MCAOĻĄ░ņŚÉņä£ Ļ░üĻ░ü 0.34 ┬▒ 0.04ņÖĆ 0.69 ┬▒ 0.06ņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż(Fig. 4B).
Ļ│Āņ░░
ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņŗĀĻ▓ĮĒĢÖņĀü Ļ▓░ņåÉ ņĀÉņłś Ļ▓Ćņé¼ņÖĆ ņĮöļäł ĒģīņŖżĒŖĖļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ MCAOĻ░Ć ņŗ¼Ļ░üĒĢ£ ņŗĀĻ▓ĮĒĢÖņĀü Ē¢ēļÅÖ ņןņĢĀļź╝ ņØ╝ņ£╝Ēé©ļŗżļŖö Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆĻ│Ā, chlorogenic acidņØś Ēł¼ņŚ¼ļŖö MCAO ņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ Ē¢ēļÅÖ ņןņĢĀļź╝ Ļ░£ņäĀņŗ£ņ╝░ļŗż. MCAO ņåÉņāüņØĆ ņØĖņ¦Ć ĻĖ░ļŖź ļ░Å ļ░śļīĆņ¬Į ņÜ┤ļÅÖņØś ņåÉņāüņØä ĒżĒĢ©ĒĢ£ Ē¢ēļÅÖ ņןņĢĀņÖĆ ļīĆļćīĒö╝ņ¦łņØś ņĪ░ņ¦üļ│æļ”¼ĒĢÖņĀü ļ│ĆĒÖöļź╝ ņØ╝ņ£╝Ēé©ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳Ļ│Ā, ņĢ×ņäĀ ņŚ░ĻĄ¼ņŚÉņä£ ņÜ░ļ”¼ļŖö ļŗżņ¢æĒĢ£ ņŗĀĻ▓ĮĒ¢ēļÅÖĒĢÖņĀü ĒģīņŖżĒŖĖņÖĆ ņĪ░ņ¦üĒĢÖņĀü ņĪ░ņé¼ļź╝ ĒåĄĒĢ┤ ĒŚłĒśłņä▒ ļćīņĪĖņżæ ļÅÖļ¼╝ļ¬©ļŹĖņŚÉņä£ chlorogenic acidņØś ņŗĀĻ▓Įļ│┤ĒśĖ ĒÜ©Ļ│╝ļź╝ ļ│┤Ļ│ĀĒĢśņśĆļŗż[26]. Chlorogenic acid Ēł¼ņŚ¼ļŖö MCAO ņåÉņāüņŚÉ ņ£ĀļÅäļÉ£ ņŗĀĻ▓ĮĒ¢ēļÅÖņØś ĻĖ░ļŖźņĀü Ļ▓░ĒĢ©ņØä ņÖäĒÖöņŗ£ņ╝░Ļ│Ā, ļćīĻ▓ĮņāēņØś ļČĆĒö╝ļź╝ Ļ░Éņåīņŗ£ņ╝░ņ£╝ļ®░, ļīĆļćīĒö╝ņ¦łņØś ņĪ░ņ¦üļ│æļ”¼ĒĢÖņĀü ļ│ĆĒÖöņÖĆ ņäĖĒżņé¼ļ®ĖņØś ņĀĢļÅäļź╝ Ļ░£ņäĀņŗ£ņ╝£, ĒŚłĒśł ņåÉņāüņ£╝ļĪ£ļČĆĒä░ ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢśņśĆļŗż[26]. ļśÉĒĢ£, chlorogenic acidņØś Ēł¼ņŚ¼ļŖö ļīĆļćīĒŚłĒśłļĪ£ ņ£ĀļÅäļÉ£ ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżļź╝ Ļ░Éņåīņŗ£ņ╝£ MCAO ņåÉņāüņ£╝ļĪ£ļČĆĒä░ ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢśņśĆļŗż[13]. ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżļŖö ļćīņåÉņāüņØś ņŻ╝ņÜö ņøÉņØĖņØ┤ļ®░, ĒŖ╣Ē׳ ņŗĀĻ▓ĮņäĖĒżļŖö ļåÆņØĆ ļīĆņé¼ņ£©Ļ│╝ ĒĢŁņé░ĒÖö ļ░®ņ¢┤ņ▓┤Ļ│äĻ░Ć ļ»ĖņĢĮĒĢśņŚ¼ ņé░ĒÖöņĀü ņŖżĒŖĖļĀłņŖżņŚÉ ļ¦żņÜ░ ņĘ©ņĢĮĒĢśļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[27,28]. ņÜ░ļ”¼ļŖö ļćīņĪĖņżæ ļ¬©ļŹĖņŚÉņä£ chlorogenic acidņØś Ēł¼ņŚ¼Ļ░Ć ļ»ĖņäĖņĢäĻĄÉņäĖĒżņØś ĒÖ£ņä▒ĒÖöļź╝ ņ¢ĄņĀ£ĒĢśĻ│Ā, pro-inflammtory ļŗ©ļ░▒ņ¦łņØĖ nuclear factor kappa B, interleukin-1╬▓, tumor necrosis factor ╬▒ļź╝ ņ¢ĄņĀ£ĒĢśņŚ¼ ĒĢŁņŚ╝ņ”ØĒÜ©Ļ│╝ļź╝ ļéśĒāĆļāäņØä ļ│┤ņŚ¼ņŻ╝ņŚłļŗż[14]. ļśÉĒĢ£, ļīĆļćīĒŚłĒśł ņåÉņāü ņŗ£ chlorogenic acidņØś ņŗĀĻ▓Įļ│┤ĒśĖ ņ×æņÜ®ņØĆ ļŗżņ¢æĒĢ£ ļŗ©ļ░▒ņ¦łņØś ņĪ░ņĀłĻ│╝ņĀĢņØä ĒåĄĒĢśņŚ¼ ņØ┤ļŻ©ņ¢┤ņ¦ł Ļ▓āņ£╝ļĪ£ ņČöņĖĪĒĢśņŚ¼, chlorogenic acidņŚÉ ņØśĒĢ┤ ļ│ĆĒÖöļÉśļŖö ļŗ©ļ░▒ņ¦łļōżņØä ĒöäļĪ£Ēģīņśżļ»╣ņŖż ļČäņäØļ▓Ģņ£╝ļĪ£ ĒāÉņāēĒĢśņśĆļŗż. ĒāÉņāēļÉ£ ļŗ©ļ░▒ņ¦ł ņżæ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ņŗĀĻ▓ĮņäĖĒżņØś ļŗżņ¢æĒĢ£ ņäĖĒż ĒÖ£ļÅÖņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ╬│-enolase ļ░£ĒśäņØś ļ│ĆĒÖöņŚÉ ņżæņĀÉņØä ļæÉņŚłļŗż. MCAO ņåÉņāüņØĆ ╬│-enolaseņØś ļ░£ĒśäņØä Ļ░Éņåīņŗ£ņ╝░Ļ│Ā, chlorogenic acid Ēł¼ņŚ¼ļŖö ņØ┤ļ¤¼ĒĢ£ Ļ░Éņåīļź╝ ņÖäĒÖöņŗ£Ēé©ļŗżļŖö Ļ▓āņØä ļ│Ė ņŚ░ĻĄ¼ļź╝ ĒåĄĒĢ┤ ĒÖĢņØĖĒĢśņśĆļŗż.
╬│-EnolaseļŖö ņŗĀĻ▓ĮļÅīĻĖ░ņØś ņ×¼ņāØĻ│╝ ņŗĀĻ▓ĮņäĖĒżņØś ņāØņĪ┤ņŚÉ Ļ┤ĆņŚ¼ĒĢśļ®░, ņŗĀĻ▓Įļ│┤ĒśĖ ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ņŚłļŗż[18,19]. ļśÉĒĢ£, ╬│-enolaseļŖö PI3K/AktņÖĆ MAPK/ERK ņŗĀĒśĖņĀäļŗ¼Ļ▓ĮļĪ£ņØś ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ņŗĀĻ▓ĮņäĖĒżņØś ņ£Āņ¦ĆņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņ¢┤, ņåÉņāüņ£╝ļĪ£ļČĆĒä░ ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢśļŖö ļŹ░ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢ£ļŗż[29]. Ļ│╝ļ░£ĒśäļÉ£ ╬│-enolaseļŖö ņŗĀĻ▓ĮņäĖĒżņØś ņä▒ņןņØä ņ┤ēņ¦äĒĢśĻ│Ā ņČĢņéŁļÅīĻĖ░ņØś ņ×¼ņāØņŚÉ Ļ┤ĆņŚ¼ĒĢśņŚ¼ ņŗĀĻ▓ĮņäĖĒżņØś ļ│┤ĒśĖņĀ£ļĪ£ņä£ ņ×æņÜ®ĒĢśņśĆļŗż[19,30]. ļö░ļØ╝ņä£, ĒĢ┤ļŗ╣Ļ│╝ņĀĢņŚÉņä£ ņŚÉļäłņ¦Ć ņāØņä▒ņŚÉ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢśļŖö ╬│-enolaseņØś Ļ░ÉņåīļŖö ņŗĀĻ▓ĮņäĖĒżņØś ņŚÉļäłņ¦Ć ļīĆņé¼Ļ│╝ņĀĢņØä ļ░®ĒĢ┤ĒĢ£ļŗżĻ│Ā ļ│╝ ņłś ņ׳ļŗż. ņÜ░ļ”¼ļŖö ņĢ×ņä£ ļīĆļćīĒŚłĒśłĻ│╝ ĻĖĆļŻ©ĒāĆļ®öņØ┤ĒŖĖ Ēł¼ņŚ¼ļĪ£ ņ£ĀļÅäļÉ£ ņŗĀņāØņĢäņØś ļīĆļćīĒö╝ņ¦ł ņåÉņāü ņŗ£ ╬│-enolaseņØś ļ░£Ēśä Ļ░Éņåīļź╝ ļ│┤Ļ│ĀĒĢśņśĆļŗż[31,32]. ļśÉĒĢ£, ļ░░ņ¢æļÉ£ ĒĢ┤ļ¦łņŗĀĻ▓ĮņäĖĒżņŚÉņä£ ĻĖĆļŻ©ĒāĆļ®öņØ┤ĒŖĖ ņ▓śļ”¼ļŖö ņŗĀĻ▓ĮņäĖĒżņØś ņé¼ļ®ĖņØä ņ£ĀļÅäĒ¢łĻ│Ā, ╬│-enolaseņØś ļ░£ĒśäņØä ĒśäņĀĆĒ׳ Ļ░Éņåīņŗ£ņ╝░ļŗż. ņØ┤ ņŚ░ĻĄ¼ļōżņØĆ ĒŚłĒśłĻ│╝ ļÅģņä▒ļ¼╝ņ¦łļĪ£ ņØĖĒĢ£ ╬│-enolaseņØś ļ░£Ēśä Ļ░ÉņåīĻ░Ć ņŗĀĻ▓ĮņäĖĒżņØś ņåÉņāüņØä ņ£ĀļÅäĒĢ©ņØä ļéśĒāĆļé┤ņŚłļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö ĒöäļĪ£Ēģīņśżļ»╣ņŖż ļČäņäØļ▓Ģņ£╝ļĪ£ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolaseņØś ļ│ĆĒÖöļź╝ ņØ╝ņ░©ņĀüņ£╝ļĪ£ ĒÖĢņØĖĒĢśņśĆĻ│Ā, reverse transverse-PCRĻ│╝ Western blot ļČäņäØņØä ĒåĄĒĢ┤ ļ¬ģĒÖĢĒĢśĻ▓ī ĒÖĢņØĖĒĢśņśĆļŗż. MCAO ņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ ļīĆļćīĒŚłĒśł ļ¬©ļŹĖņŚÉņä£ ╬│-enolaseņØś ļ░£Ēśä Ļ░ÉņåīņÖĆ chlorogenic acidņØś Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolase ļ░£ĒśäņØś Ļ░Éņåī ņÖäĒÖöļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. ņĄ£ĻĘ╝, ņÜ░ļ”¼ļŖö chlorogenic acidĻ░Ć ļīĆļćīĒŚłĒśł ņåÉņāüņ£╝ļĪ£ ņ£ĀļÅäļÉ£ ņØĖņé░ĒÖöļÉ£ AktņÖĆ ņØĖņé░ĒÖöļÉ£ BadņØś ļ░£Ēśä ņłśņżĆņØś Ļ░Éņåīļź╝ ņÖäĒÖöņŗ£ņ╝░ņ£╝ļ®░, ņØĖņé░ĒÖöļÉ£ BadņÖĆ 14-3-3 ļŗ©ļ░▒ņ¦łņØś Ļ▓░ĒĢ®ļĀźņØś Ļ░Éņåīļź╝ Ļ░£ņäĀņŗ£ņ╝░Ļ│Ā, ņäĖĒżņé¼ļ®ĖņØś ĻĖ░ņĀäņØä ņ¢ĄņĀ£ĒĢ©ņØä ļ│┤Ļ│ĀĒĢśņśĆļŗż[33]. ņØ┤ļōż Ļ▓░Ļ│╝ļŖö chlorogenic acidĻ░Ć PI3K/Akt ņŗĀĒśĖņĀäļŗ¼ņ▓┤Ļ│äļź╝ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ļīĆļćīĒŚłĒśłļĪ£ ņØĖĒĢ£ ņåÉņāü ņŗ£ ņŗĀĻ▓ĮņäĖĒżļź╝ ļ│┤ĒśĖĒĢ©ņØä ļéśĒāĆļéĖļŗż. ╬│-enolaseļŖö PI3K/AktņØś ņŗĀĒśĖņĀäļŗ¼Ļ▓ĮļĪ£ņØś ĒÖ£ņä▒ĒÖöļĪ£ ņŗĀĻ▓ĮņäĖĒżņØś ņ×¼ņāØ ļ░Å ņŗĀĻ▓ĮņäĖĒżņØś ļ│┤ĒśĖņ×æņÜ®ņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖņ׳ļŗż[30]. ņØ┤ļōż ļ│┤Ļ│ĀņŚÉ ļö░ļź┤ļ®┤, chlorogenic acidĻ░Ć ╬│-enolaseļź╝ ņĪ░ņĀłĒĢśĻ│Ā PI3K/AktņØś ņŗĀĒśĖņĀäļŗ¼Ļ▓ĮļĪ£ļź╝ ĒÖ£ņä▒ĒÖöĒĢśņŚ¼ ņäĖĒżņØś ņé¼ļ®ĖņØä ņ¢ĄņĀ£ĒĢ©ņ£╝ļĪ£ņŹ© ņŗĀĻ▓ĮņäĖĒżņØś ņāØņĪ┤ ļ░Å ļ│┤ĒśĖ ĻĖ░ļŖźņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗżĻ│Ā ļ│╝ ņłś ņ׳ļŗż. ļö░ļØ╝ņä£ chlorogenic acidņØĆ MCAOņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉśļŖö ĒŚłĒśłņä▒ ņåÉņāüņŚÉ ļīĆĒĢ┤ ╬│-enolase ļ░£ĒśäņØä ņ£Āņ¦ĆĒĢśļ®░, ņØ┤ļ¤¼ĒĢ£ ╬│-enolase ļ░£ĒśäņØś ņ£Āņ¦ĆļŖö ņŗĀĻ▓ĮņäĖĒżņØś ļ│┤ĒśĖņ×æņÜ®ņŚÉ Ļ┤ĆņŚ¼ĒĢ£ļŗż. Chlorogenic acidļŖö ĒĢŁņé░ĒÖö, ĒĢŁņĢö, ĒĢŁņŚ╝ņ”Ø ļō▒ ļŗżņ¢æĒĢ£ ņĢĮļ”¼ĒĢÖņĀü ĒŖ╣ņä▒ņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ņ£╝ļ®░, ņ×äņāüņĀüņ£╝ļĪ£ Ļ│ĀĒśłņĢĢ ĒÖśņ×ÉņØś ĒśłņĢĢņØä Ļ░Éņåīņŗ£ĒéżļŖö ĒÜ©Ļ│╝Ļ░Ć ņ׳ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż[34]. ļśÉĒĢ£, chlorogenic acidļŖö ļÅģņä▒ņ£╝ļĪ£ ņØĖĒĢ£ ņŗ¼Ļ░üĒĢ£ ļČĆņ×æņÜ®ņØä ņØ╝ņ£╝Ēéżņ¦Ć ņĢŖĻĖ░ ļĢīļ¼ĖņŚÉ ņĢłņĀäĒĢśĻ│Ā ĒÜ©Ļ│╝ņĀüņØĖ ļ¼╝ņ¦łļĪ£ ļ│┤Ļ│ĀļÉśņ¢┤, ņ×äņāüņĀü ņØæņÜ®ņØä ņ£äĒĢ┤ ņżæņÜöĒĢ£ ļ¼╝ņ¦łļĪ£ Ļ░äņŻ╝ļÉśĻ│Ā ņ׳ļŗż[35]. ņØ┤ļōż ņŚ░ĻĄ¼ļōżņØä ĒåĀļīĆļĪ£ chlorogenic acidļŖö ļćīĒŚłĒśłņØä ĒżĒĢ©ĒĢ£ ņŚ╝ņ”Øņä▒ ņŗĀĻ▓Įņ¦łĒÖśņØś ņ╣śļŻīņŚÉļÅä ņĀüņÜ® Ļ░ĆļŖźĒĢ£ ļ¼╝ņ¦łļĪ£ ĒīÉļŗ©ļÉ£ļŗż. Chlorogenic acidņŚÉ ņØśĒĢ£ ╬│-enolaseņØś ņĪ░ņĀł ĻĖ░ņĀäņØĆ ņĢäņ¦ü ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ņ¦Ć ņĢŖņ¦Ćļ¦ī, MCAO ņåÉņāü ņŗ£ chlorogenic acid Ēł¼ņŚ¼ņŚÉ ņØśĒĢ£ ╬│-enolase ļ░£Ēśä Ļ░ÉņåīņØś ņÖäĒÖöĻ░Ć ņŗĀĻ▓Įļ│┤ĒśĖ ĒÜ©Ļ│╝ņŚÉ Ļ┤ĆņŚ¼ĒĢ©ņ£╝ļĪ£, ļ│Ė ņŚ░ĻĄ¼ņØś Ļ▓░Ļ│╝ļŖö chlorogenic acidņØś ļćīņ¦łĒÖś ņ╣śļŻīņĀ£ļĪ£ņä£ņØś ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ņĀ£ņŗ£ĒĢĀ ņłś ņ׳ļŗż. Ļ▓░ļĪĀņĀüņ£╝ļĪ£, ļ│Ė ņŚ░ĻĄ¼ļŖö chlorogenic acidĻ░Ć MCAOļĪ£ ņ£ĀļÅäļÉ£ ļīĆļćīĒŚłĒśł ņŗ£ ╬│-enolaseņØś ļ░£ĒśäņØä ņĪ░ņĀłĒĢśņŚ¼ ņŗĀĻ▓ĮņäĖĒżņØś ļ│┤ĒśĖņŚÉ Ļ┤ĆņŚ¼ĒĢ©ņØä ĻĘ£ļ¬ģĒĢśņśĆļŗż.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print